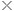|
土壤、植物和大气中由植物蒸腾耗水引起水流运移而构成的连续体。由于此系统内的水流及其水势均能保持连续状态,所以澳大利亚学者J.R.菲利普将其命名为土壤—植物—大气连续体(The Soil-Plant-Atmosphere Continuum),简称SPAC。大气和土壤是植物赖以生存的环境,通过对SPAC的研究,可揭示植物与环境之间的水分传输规律,为进行田间水、热平衡,研究节水机理,确定植物需水量及制定灌溉排水计划提供理论根据。 水流传输 在土壤—植物—大气系统中,土壤水作为水流的“源”,大气是“汇”,植物是二者之间的导水“介质”。植物蒸腾时,水流首先从靠近根系的土壤中向根表皮组织移动,依次通过根的表皮、皮层、内皮层、中柱鞘及其中柱薄壁组织到达木质部中的导管或管胞,再沿茎杆流到叶部组织,最后在大气蒸发力作用下,液态水转化为气态水并由气孔或角质层逸出,扩散到大气中。在高等植物中,根部和叶部均为活细胞,而介于根、叶之间的木质部则为死细胞。植物与环境发生水流联系时,植物能依靠本身的活细胞对其体内的水流起到一定控制作用。这是该系统与不受生物控制的一般水流系统的主要差别。在大气蒸发条件下,植物的根系吸水速率与叶面蒸腾速率总是处于动平衡状态。只有当根系吸水速率基本满足叶面蒸腾的需要时,植物才得以正常生长,反之将出现植物缺水现象,甚至凋萎或死亡。 植物叶部气孔既是散失水分的主要“门窗”,又是吸收大气中二氧化碳的通道。植物水分亏缺时关闭气孔可降低蒸腾速率,但同时也降低了二氧化碳的吸入速率,抑制了光合作用。人们可在一定程度上调控土壤、大气水分。合理调节控制土壤—植物—大气系统的水流,对于提高植物活力、获取最高生物产量有着直接影响。

土壤—植物—大气系统中水势分布示意图
水势 该系统中的水,无论是液态或气态,均具有一定的“能”(即做功的能力)。这种“能”的相对值即为水势。通常水势是相对于某一位置上的纯自由水而言的。该系统中不同位置之间水势的差值称为水势差。水势差能驱使水克服摩阻力和重力,由水势高处向水势低处流动。叶面产生蒸腾时,首先引起叶水势下降并使叶、茎之间出现水势差和水势梯度,从而使水由茎部流向叶部;接着在茎、根间和根、土间发生连锁反应,最终形成由土壤经植物到大气的水流。土壤—植物—大气系统中的总势差可达到几十个兆帕,在干燥气候下甚至超过100 MPa。由叶面到大气这一段,由于水发生相变(即由液态转变为气态),能量消耗最大,所以水势差也最大,占总势差的绝大部分。从土壤到植物及在植物内部的水势差均较小,一般只有几个兆帕。 土壤—植物—大气系统中水势分布与蒸腾的大致关系如图所示。图中曲线1、2和曲线3、4,分别表示土壤水势较高和较低的2种情况;曲线1、3所代表的蒸腾速率,分别比曲线2和4低。 水流阻力 沿水流方向的每一部位或区段,如根周围土壤、根部、叶气孔等,都对系统内的水流产生阻力。SPAC中的水流阻力有2种:一种是关于液态水的水力阻力;另一种是关于气态水的扩散阻力。 液态水流服从达西定律,由此可导出水力阻力,即

式中,Rh为水力阻力,表示产生单位水流通量所需要的水势差;ΔΨ为水势差;q为水流通量,即单位过水断面面积上通过的流量。 气态水流服从菲克定律,由此可导出扩散阻力,即

式中,RD为扩散阻力,表示产生单位水流通量所需的水汽浓度差;Δc为水汽浓度差;q的意义同式(1)。 显然,这2种水流阻力具有不同的物理意义,因而不能互比,也不能叠加。 水流模型 当蒸腾速率和植物体内的水分贮量均无变化时,即流经各段的水流通量相等时,该系统的水流属于稳定流。美国著名土壤物理学家W.R.加德纳对这种稳定流提出如下的数学模型:

式中,E0、RS、ΨL分别为潜在蒸腾速率、气孔阻力和叶水势;函数f(E0,RS,ΨL)为流向大气的实际蒸腾速率;ΨR、ΨS分别为根部水势和土壤水势;RP为植物体内的水力阻力;K为土壤水力传导度;L为单位土体内有效根的总长度。 土壤—植物—大气系统中的水流现象十分复杂,目前对其中许多理论问题,如连续性、根系吸水机理等尚未研究透彻,某些重要参数(如阻力、扩散度等)的测定还缺乏完备的测试技术。因此,现有各种描述这一系统的数学模型还不能普遍应用和推广。
|